非编码 RNA(non-coding RNA,ncRNA)指不(或主要不)以直接编码蛋白质为功能的 RNA 分子。基因组转录产物中,ncRNA 在数量与调控复杂度上往往不亚于 信使 RNA(messenger RNA,mRNA)。理解 ncRNA 的分类逻辑——按长度、亚细胞定位、生物发生通路还是分子机器归属——是阅读 RNA 干扰(RNA interference,RNAi)、小干扰 RNA(small interfering RNA,siRNA)药物、RNA 测序(RNA sequencing,RNA-seq)与 RNA 基础模型 文献的共同前置。
段末注释:ncRNA 与「非蛋白编码转录本」在组学语境下常混用,但严格说部分 lncRNA 可含短 开放阅读框(open reading frame,ORF)并产生微肽;是否算「编码」取决于定义边界。
读前说明
- 本文按「管家型 ncRNA → 小调控 RNA → 长调控 RNA → 衍生/情境型 RNA」组织,并非穷尽所有已命名亚型(如 tiRNA、sdRNA、vault RNA 等仅在表中点名)。
- 长度界限常用 200 核苷酸(nucleotide,nt)区分 长非编码 RNA(long non-coding RNA,lncRNA)与 小 ncRNA(small ncRNA,sncRNA),但 核糖体 RNA(ribosomal RNA,rRNA)、转运 RNA(transfer RNA,tRNA)等可远超 200 nt 仍不归入 lncRNA——分类需结合功能而非单看长度。
- 数学上长度分布、配对概率等 occasionally 用 $L$ 表示序列长度、$\Delta G$ 表示折叠自由能;下文以生物学描述为主。
- 配图均为科普示意(PNG),非某篇论文原图;细节以最新综述与数据库(Rfam、RNAcentral、GENCODE)为准。
- 插图路径:资源位于与本文同名的目录
siRNA-00-前置知识-02-ncRNA系统综述/;站点已开启post_asset_folder与 hexo-asset-image,正文图链仅写文件名(如fig01-ncRNA-taxonomy.png)。编辑器内 Markdown 预览可能不显示,请以hexo server或部署后页面为准。
1. 什么是 ncRNA:与 mRNA 的边界
1.1 定义与基因组视角
ncRNA 来自基因组 DNA 转录,但成熟产物不作为典型 mRNA 被 核糖体 翻译成全长蛋白。人类基因组中,蛋白质编码基因仅占约 1–2%,而 非编码转录 占转录本的大多数;其中一部分是 技术噪音 或 低水平转录,另一部分具有明确的 结构、催化 或 调控 功能。
mRNA 与 ncRNA 的区分维度包括:
| 维度 | mRNA | ncRNA(典型) |
|---|---|---|
| 主要功能 | 蛋白合成模板 | 结构、催化、调控、支架 |
| 5’ 端 | 7-甲基鸟苷帽(m7G cap)常见 | 多样:有帽/无帽/三磷酸 |
| 3’ 端 | poly(A) 尾常见 | 多样:聚腺苷酸化、加工末端、环化 |
| 剪接 | 外显子–内含子剪接产生编码序列 | 剪接产生 lncRNA/circRNA 或内含子降解 |
| 保守性 | 蛋白 ORF 保守 | 高度分化:从极端保守(rRNA)到快速演化(部分 lncRNA) |
段末注释:ORF 为可能翻译成肽段的核苷酸阅读框;lncRNA 常缺乏长 ORF 但并非绝对无肽。
1.2 分类总览
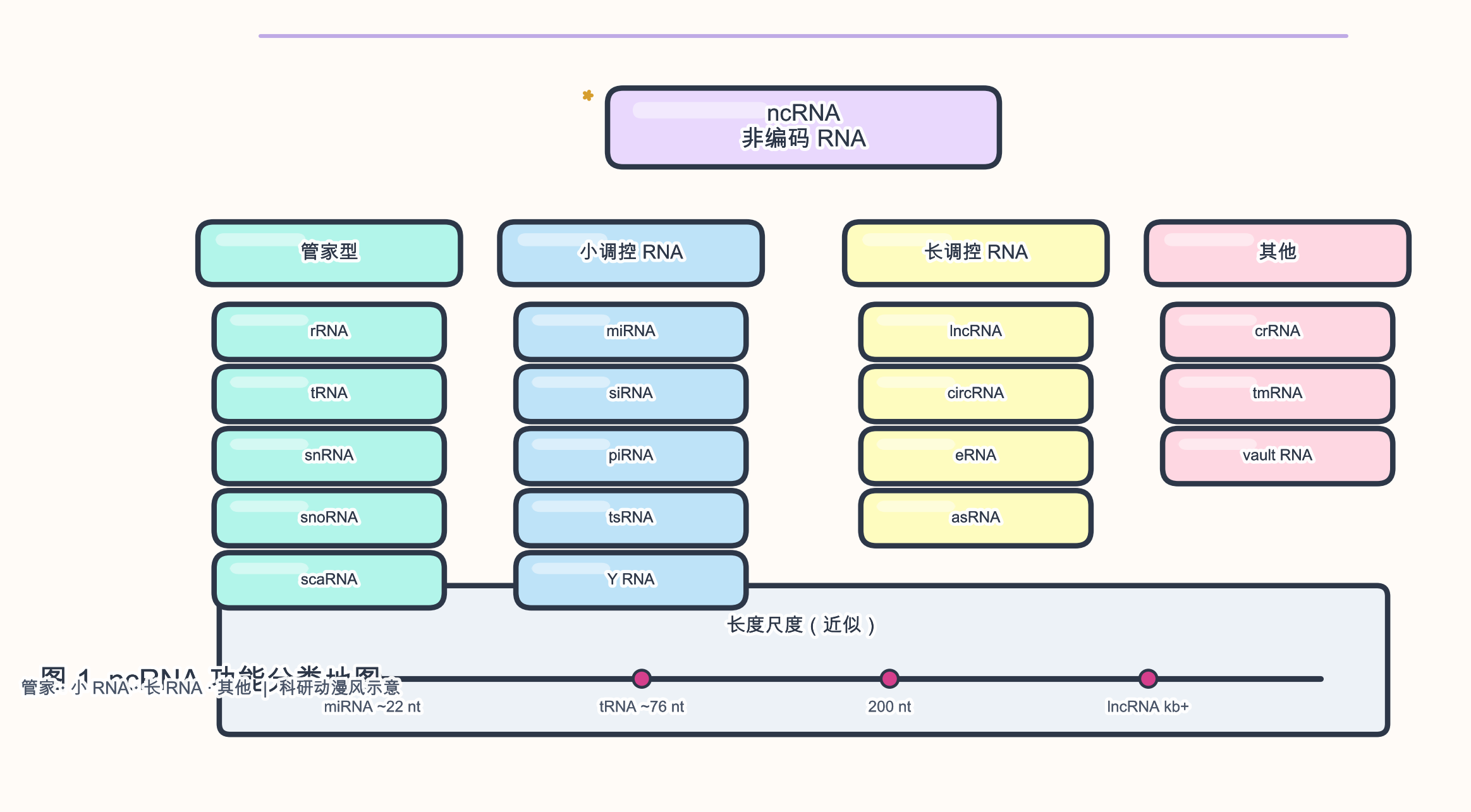
图 1 给出阅读本文的「地图」:管家型分子支撑翻译与 RNA 加工;小调控 RNA 多通过 碱基配对 与蛋白效应器发挥作用;长调控 RNA 常通过 染色质、转录、剪接 与 信号 网络整合;衍生型(如 环状 RNA、CRISPR RNA)则依赖特定生物发生或人工语境。
2. 管家型 ncRNA:翻译与 RNA 加工的核心
2.1 rRNA(核糖体 RNA)
生物学角色:rRNA 是 核糖体 的主体 RNA 成分,构成 肽酰转移酶 活性中心(核酶 性质),并搭建 mRNA、tRNA 与翻译因子的装配平台。
参与的生物学过程:翻译(translation)全过程;核糖体发生(ribosome biogenesis)在 核仁 中进行,涉及 前 rRNA(pre-rRNA)转录、切割与修饰。
序列与结构特征:
- 人类 45S 前体经加工产生 18S(小亚基)、5.8S 与 28S(大亚基);5S rRNA 由独立基因 RNA 聚合酶 III(RNA polymerase III,Pol III)转录。
- 长度:成熟 18S ~1.9 kb,28S ~5 kb 量级(物种差异大)。
- 结构:高度保守的 二级/三级结构 域,与 核糖体蛋白(ribosomal protein,r-protein)形成复杂 RNP(ribonucleoprotein)。
研究与应用:抗生素(如部分药物靶向细菌 rRNA 差异)、核糖体图谱(ribosome profiling)解析翻译调控、RNA 修饰 与疾病(核糖体病 ribosomopathy)、冷冻电镜 核糖体结构。
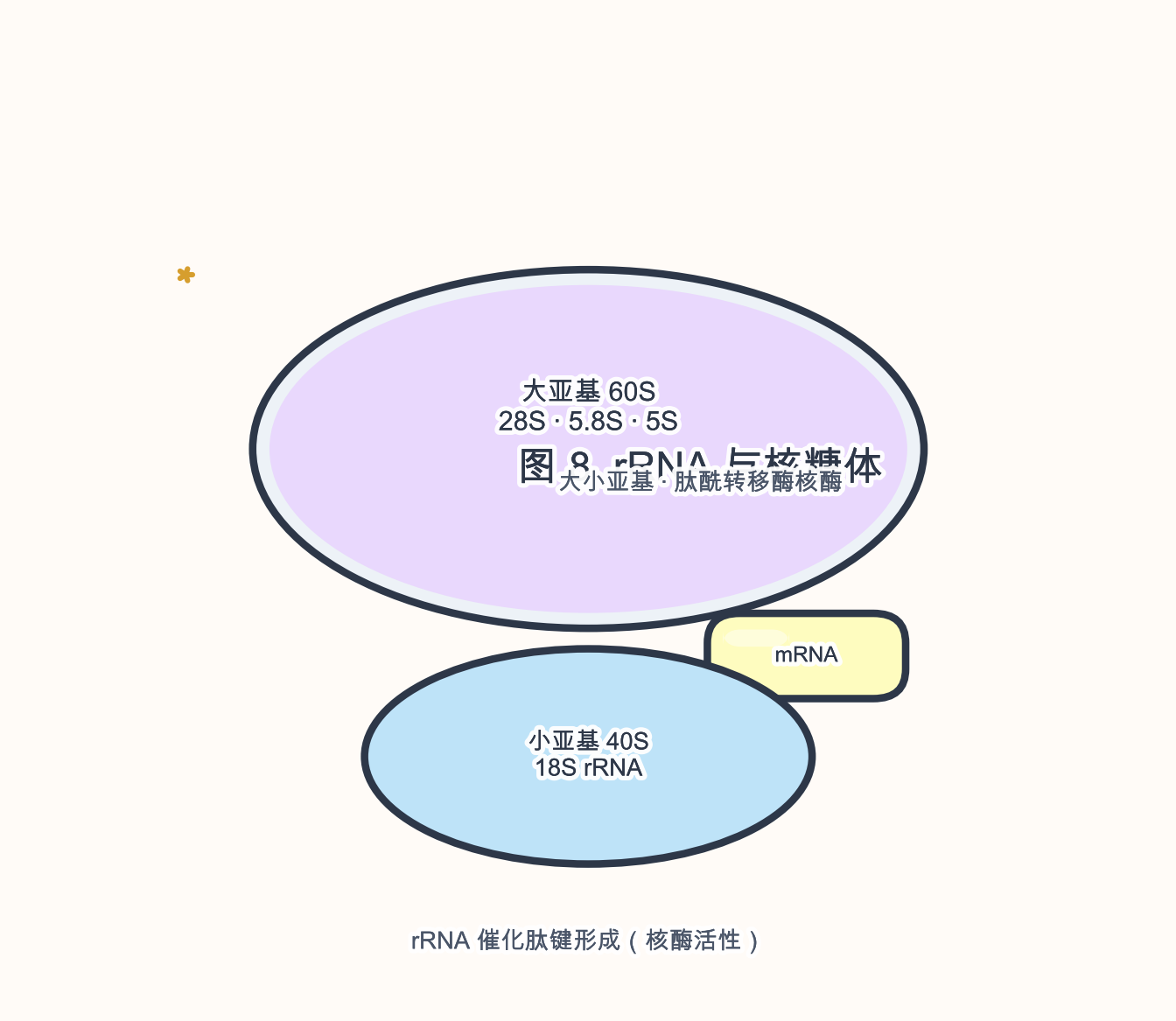
段末注释:Pol III 转录短 ncRNA(tRNA、5S rRNA 等);Pol I 主要转录 rRNA 前体。
2.2 tRNA(转运 RNA)
生物学角色:tRNA 将 氨基酸 按 mRNA 密码子 运至核糖体,是遗传密码与蛋白序列之间的「适配器」。
参与的生物学过程:翻译延伸;氨酰-tRNA 合成酶(aminoacyl-tRNA synthetase)催化 氨酰化;部分 tRNA 片段(tRF/tiRNA)进入应激调控(见 §6)。
序列与结构特征:
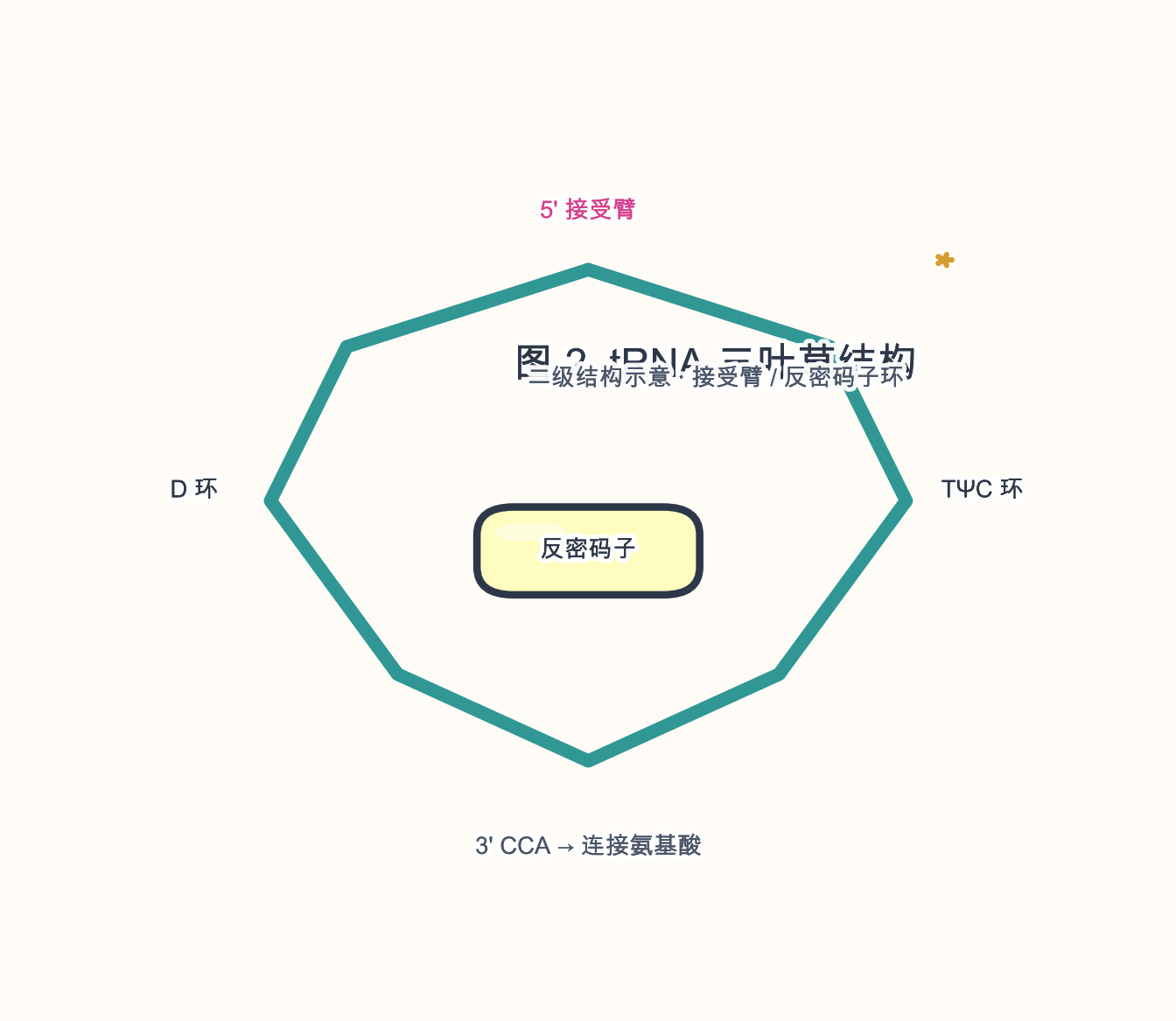
- 典型长度 ~76–90 nt(人类多数 ~73–95 nt)。
- 三叶草 二级结构:接受茎(CCA-3’ 氨酰化末端)、D 环、反密码子环、TΨC 环、可变环;三级结构呈 L 形。
- 修饰核苷酸 丰富(如 假尿苷 Ψ、m1A),影响折叠与解码。
研究与应用:tRNA 测序 监测翻译池;肿瘤与应激中 tRNA 表达与修饰重编程;合成生物学中的 正交 tRNA;tRNA 衍生小 RNA 作为生物标志物。
2.3 snRNA(小核 RNA)与剪接体
生物学角色:snRNA 是 剪接体(spliceosome)的核心组分,识别 pre-mRNA 上的 剪接位点 并催化 内含子 移除。
参与的生物学过程:pre-mRNA 剪接;U1 snRNA 识别 5’ 剪接位点,U2 snRNA 与 分支点 腺苷配对,U6 snRNA 具有 催化 活性(与 U2 协作),U5 snRNA 协调外显子连接。
序列与结构特征:
- 长度约 100–300 nt(因种类而异)。
- Sm 环 蛋白结合 Sm 位点;U6 与 U4 形成双链后在激活时解旋。
- 次要剪接体(U12-type)使用 U11/U12/U4atac/U5/U6atac 处理少数 AT–AC 内含子。
研究与应用:剪接因子 突变导致的遗传病;可变剪接 与癌症;反义寡核苷酸 调节特定外显子 inclusion(如 SMA 治疗语境下的 SMN2 外显子 7);RNA-seq 剪接分析(rMATS、LeafCutter 等)。
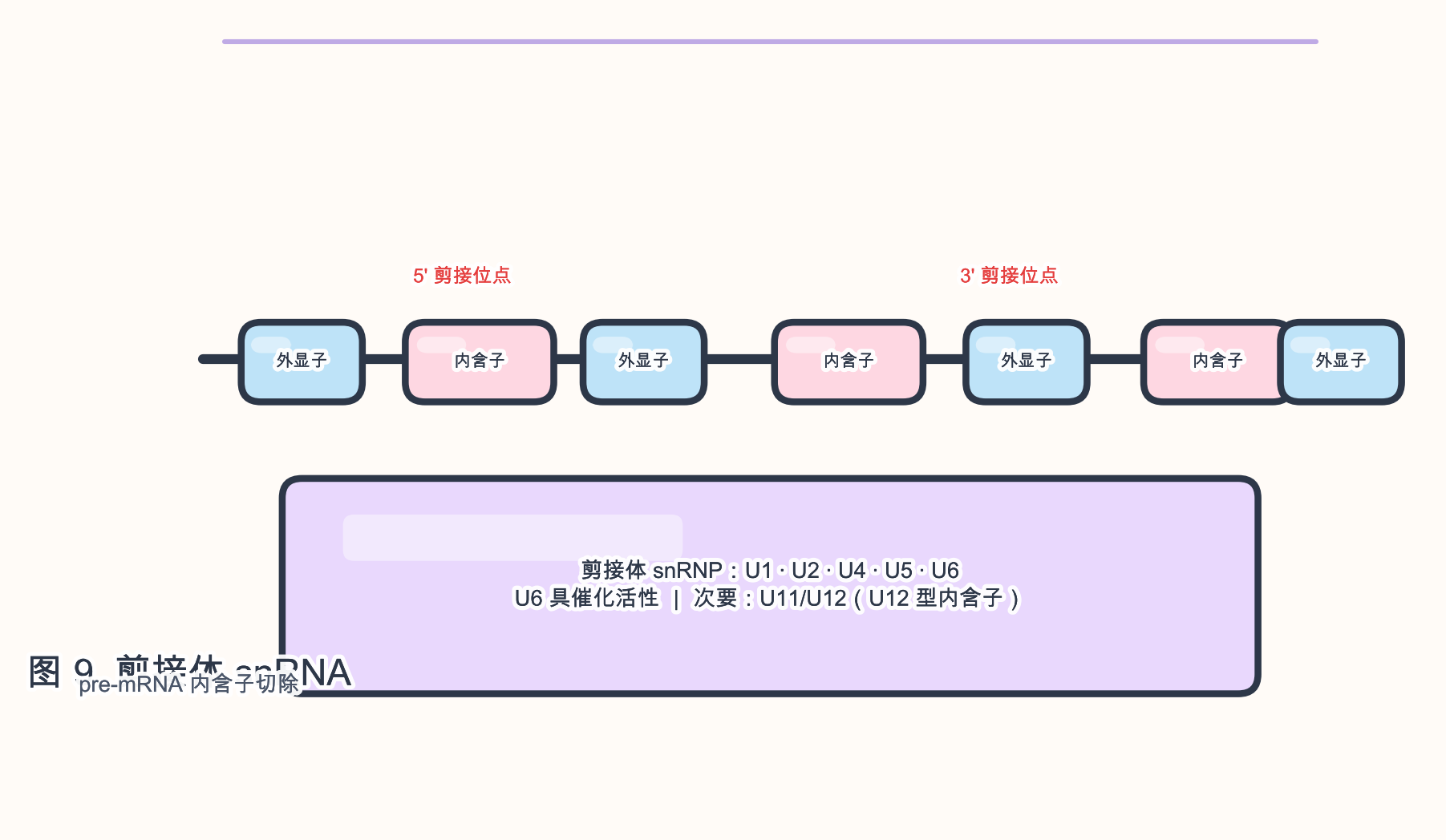
段末注释:snRNP 为 snRNA 与蛋白组成的 小核核糖核蛋白颗粒。
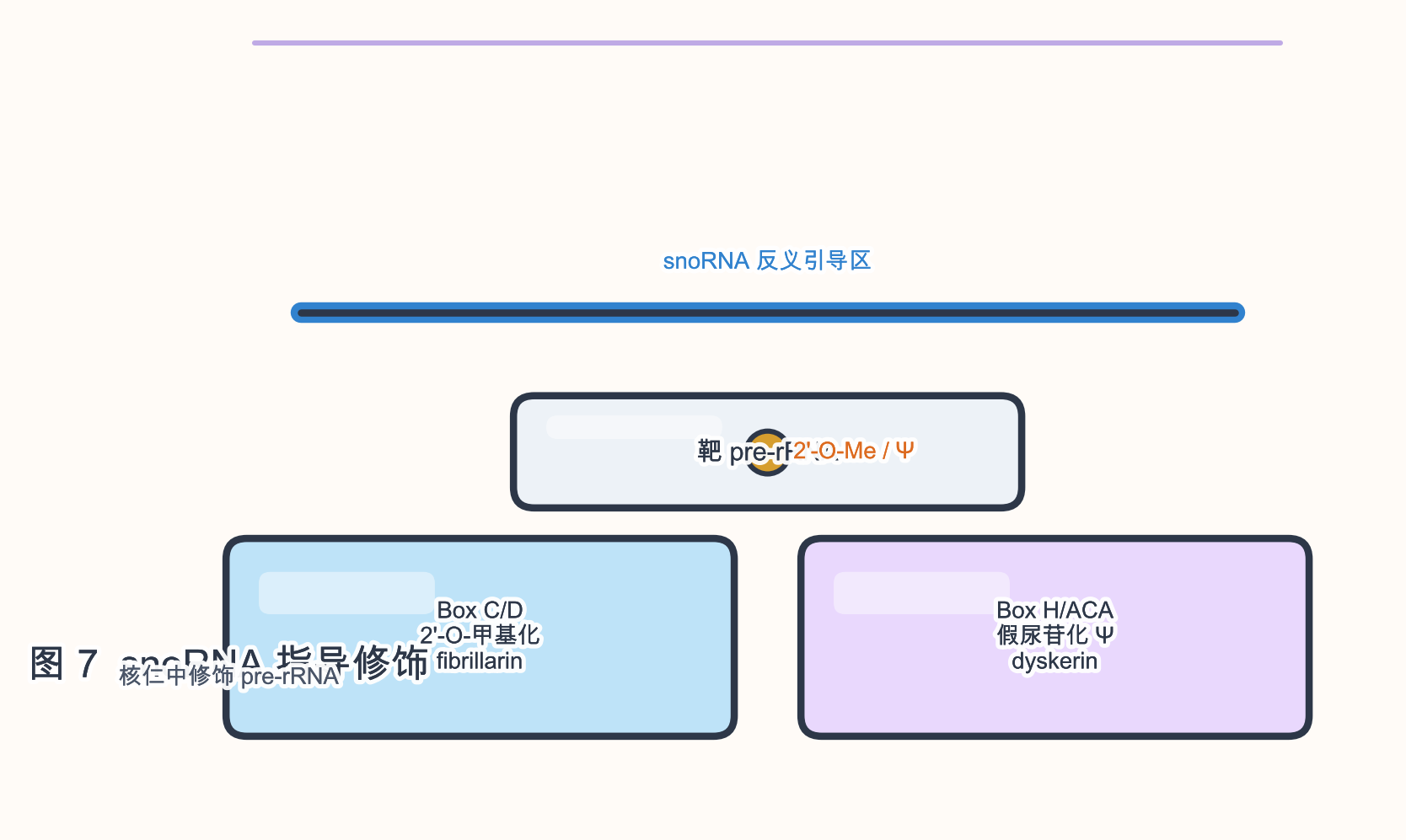
2.4 snoRNA(小核仁 RNA)与 scaRNA
生物学角色:snoRNA 在 核仁 中指导 rRNA、snRNA 等靶 RNA 的 2’-O-甲基化(box C/D)或 假尿苷化(box H/ACA)。scaRNA(small Cajal body RNA)多在 Cajal 体 中指导 snRNA 修饰。
参与的生物学过程:核糖体发生;剪接体 RNP 成熟;与 端粒酶 RNA(TERC)等 H/ACA 家族有进化联系。
序列与结构特征:
- 长度约 60–300 nt。
- box C/D:C(RUGAUGA)、D 盒及 反义引导区 与靶配对。
- box H/ACA:hairpin-hinge-hairpin-tail 结构,反义环 指定 Ψ 位点。
研究与应用:先天性角化不良 等 H/ACA 通路缺陷;snoRNA 宿主基因内含子定位;部分 snoRNA 加工产生 sdRNA 进入 miRNA 样通路(边界案例)。
3. 小调控 RNA:RNAi 与 Argonaute 家族
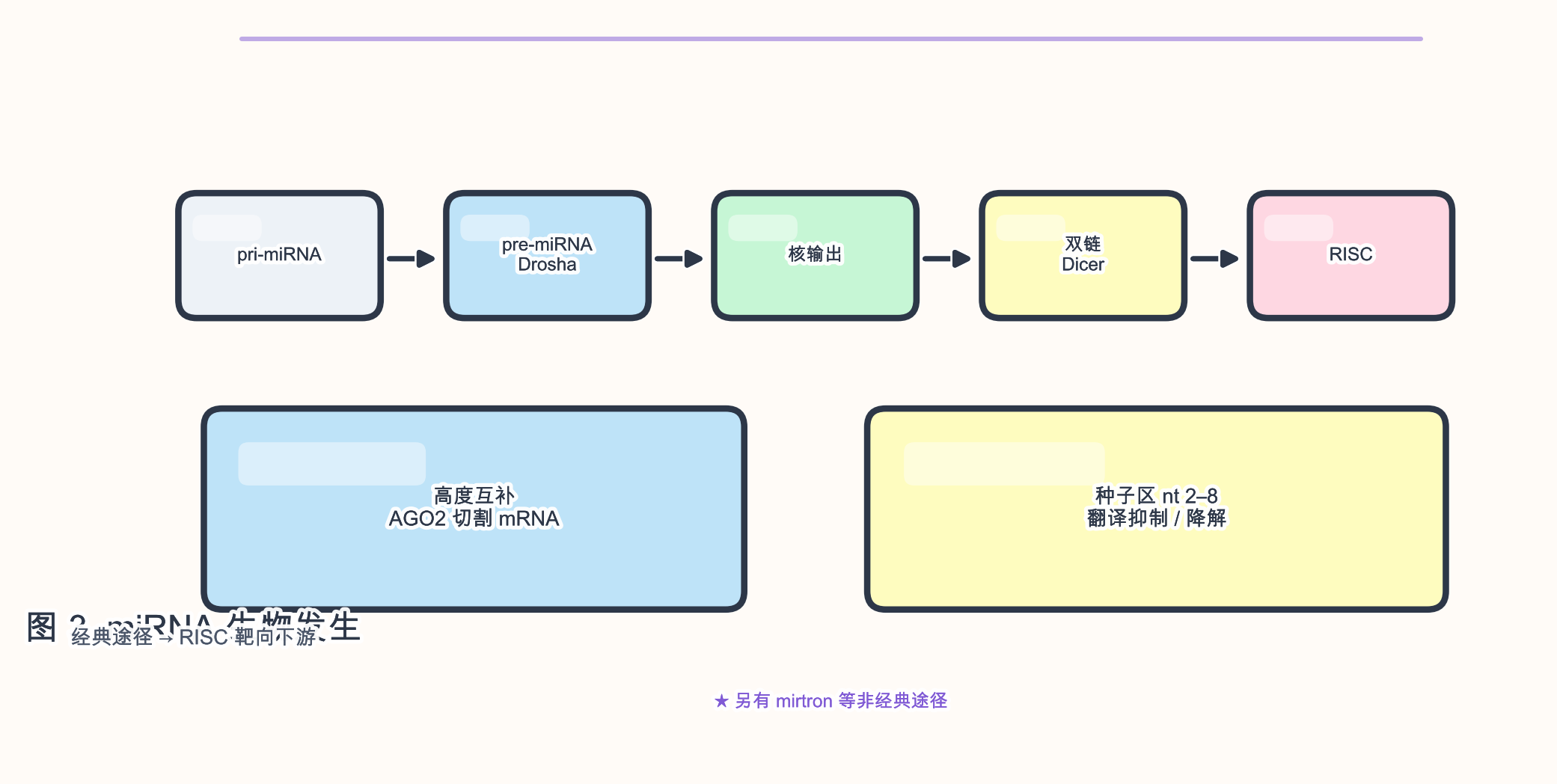
3.1 miRNA(微 RNA)
生物学角色:miRNA 是内源性 ~22 nt 单链 RNA,装载入 RNA 诱导沉默复合体(RNA-induced silencing complex,RISC),以 种子区(5’ 端约第 2–8 nt)为主识别靶 mRNA,导致 翻译抑制 与/或 mRNA 降解。
参与的生物学过程:发育、分化、免疫、代谢、肿瘤等几乎全生理路径;与 转录因子 网络形成反馈。
序列与结构特征:
- 成熟体 ~21–23 nt;前体 pre-miRNA 为 ~60–70 nt 发夹,pri-miRNA 更长。
- 5’ 端磷酸、3’ 端 常有 2’-O-甲基 修饰(HENMT1 等)。
- 靶配对:哺乳动物多为 不完全配对;植物中常见 近乎完全配对 导致 AGO 切割。
生物发生(典型途径):Pol II 转录 pri-miRNA → Drosha/DGCR8(微处理器)→ pre-miRNA 输出至胞质 → Dicer/TRBP/PACT → 双链 → AGO2 装载 → 保留 引导链。
研究与应用:miRNA 模拟物 与 抗 miRNA(antimiR,如 locked nucleic acid,LNA);液体活检 miRNA 签名;AGO2 CLIP-seq 鉴定靶标;与 siRNA 药物共享递送与化学修饰经验。
段末注释:RISC 核心含 AGO 蛋白;人 AGO2 具 slicer 切割活性。
3.2 siRNA(小干扰 RNA)
生物学角色:siRNA 通常为 ~19–25 bp 双链,引导 RISC 对靶 mRNA 进行 高度互补 依赖的 AGO2 切割,实现 基因敲低。
参与的生物学过程:外源 siRNA 多用于实验或治疗性敲低;内源性 endo-siRNA 在生殖细胞、干细胞等场景参与 转座子 与 基因 调控(与 piRNA 有交叉)。
序列与结构特征:
- 双链,常带 2 nt 3’ 突出端;5’ 磷酸 利于 RISC 装载。
- 热力学不对称 决定 引导链/过客链 选择。
- 与 miRNA 相比:更要求全链配对 才能高效切割(哺乳动物胞质外源 siRNA)。
研究与应用:见本系列 siRNA-01 至 siRNA-05;寡核苷酸药物(如肝脏 GalNAc–siRNA);植物 RNAi 植保;高通量 siRNA 文库筛选。
与 miRNA 的对照见 §7 表 1;机制示意图亦可参阅 siRNA-01-生物学机制与功能原理 配图。
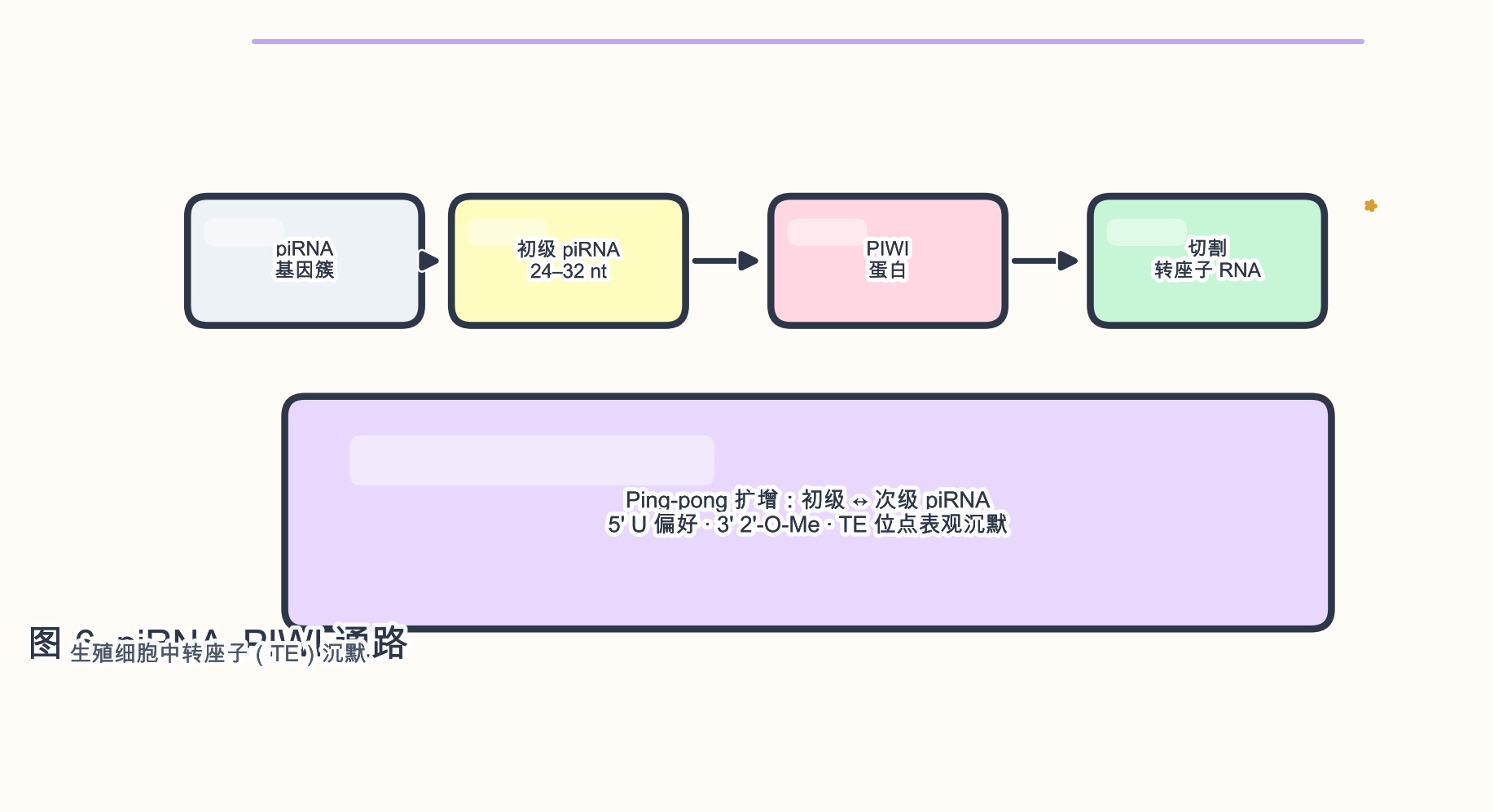
3.3 piRNA(PIWI 相互作用 RNA)
生物学角色:piRNA(~24–32 nt)与 PIWI 家族蛋白结合,在 生殖细胞 中沉默 转座子(transposable element,TE),维持基因组稳定性。
参与的生物学过程:ping-pong 扩增(primary/secondary piRNA 循环);DNA 甲基化 与 H3K9me3 等表观标记在 TE 位点沉积。
序列与结构特征:
- 5’ 尿苷 偏好(初级);3’ 端 2’-O-甲基化。
- 来源于基因组 piRNA 簇(often 单链 转录本)。
研究与应用:不育、肿瘤 生殖系 标志物、PIWI 蛋白在多种癌症中异常表达的研究;与 siRNA/miRNA 通路在 AGO 家族上分工明确。
3.4 其他小 RNA(选读)
| 类型 | 长度/来源 | 主要功能 | 研究要点 |
|---|---|---|---|
| tsRNA / tRF | tRNA 切割片段,~14–40 nt | 应激、翻译调控、免疫 | 与经典 miRNA 区分生物发生 |
| sdRNA | snoRNA 衍生 | 部分进入 RISC | 注释陷阱 |
| Y RNA | ~80–110 nt | Ro RNP、DNA 复制 相关 | 自身免疫 Sjögren 等 |
| vault RNA | ~86–141 nt | vault 颗粒,功能未完全清楚 | 药物载体研究 |
| 7SK RNA | ~330 nt | 抑制 P-TEFb,调控 Pol II 暂停 | HIV Tat、转录延伸 |
| RNase P RNA | 催化 tRNA 5’ 成熟 | 核酶 | 进化保守 |
段末注释:TE 为可移动遗传元件,piRNA 主要压制其活性。
4. 长调控 RNA 与环状 RNA
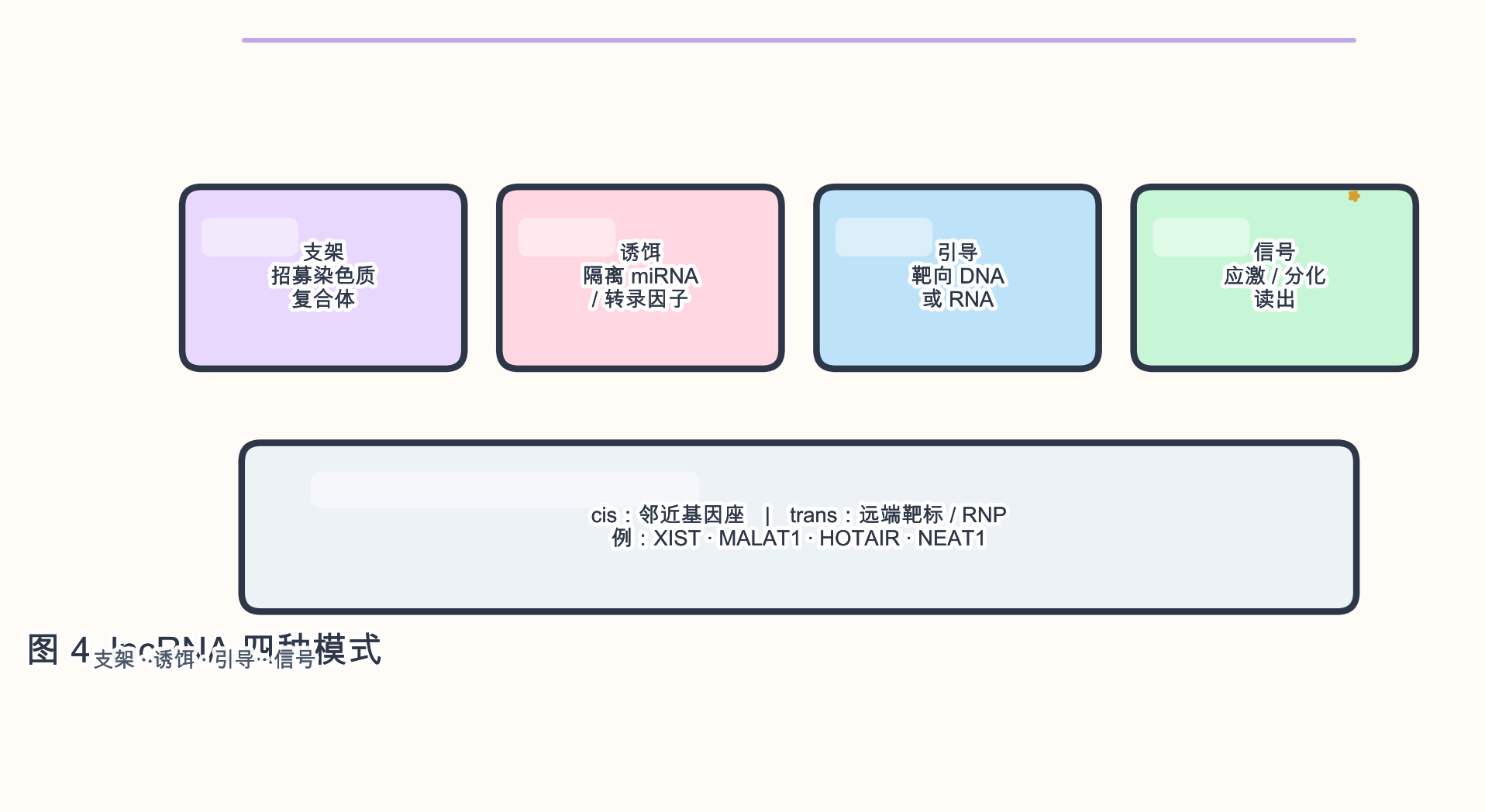
4.1 lncRNA(长非编码 RNA)
生物学角色:lncRNA(通常 >200 nt,无长 ORF)通过 支架、诱饵、引导、信号 等模式调节 染色质、转录、剪接、蛋白定位 与 信号通路。
参与的生物学过程:X 染色体失活(XIST)、核旁斑(NEAT1–MALAT1 网络)、胚胎发育、肿瘤转移(HOTAIR 等)。
序列与结构特征:
- 长度 数百 nt 至 >100 kb;剪接、聚腺苷酸化 常见。
- 序列保守性 常低于 蛋白编码基因,但 局部结构域 与 重复元件 可保守。
- 亚细胞定位 决定功能:核(染色质)、胞质(miRNA 海绵)、膜关联 等。
研究与应用:lncRNA 靶向 反义寡核苷酸;CRISPRi/CRISPRa 表观编辑;癌症 预后签名;RNA 结合蛋白(RNA-binding protein,RBP)图谱(CLIP)。
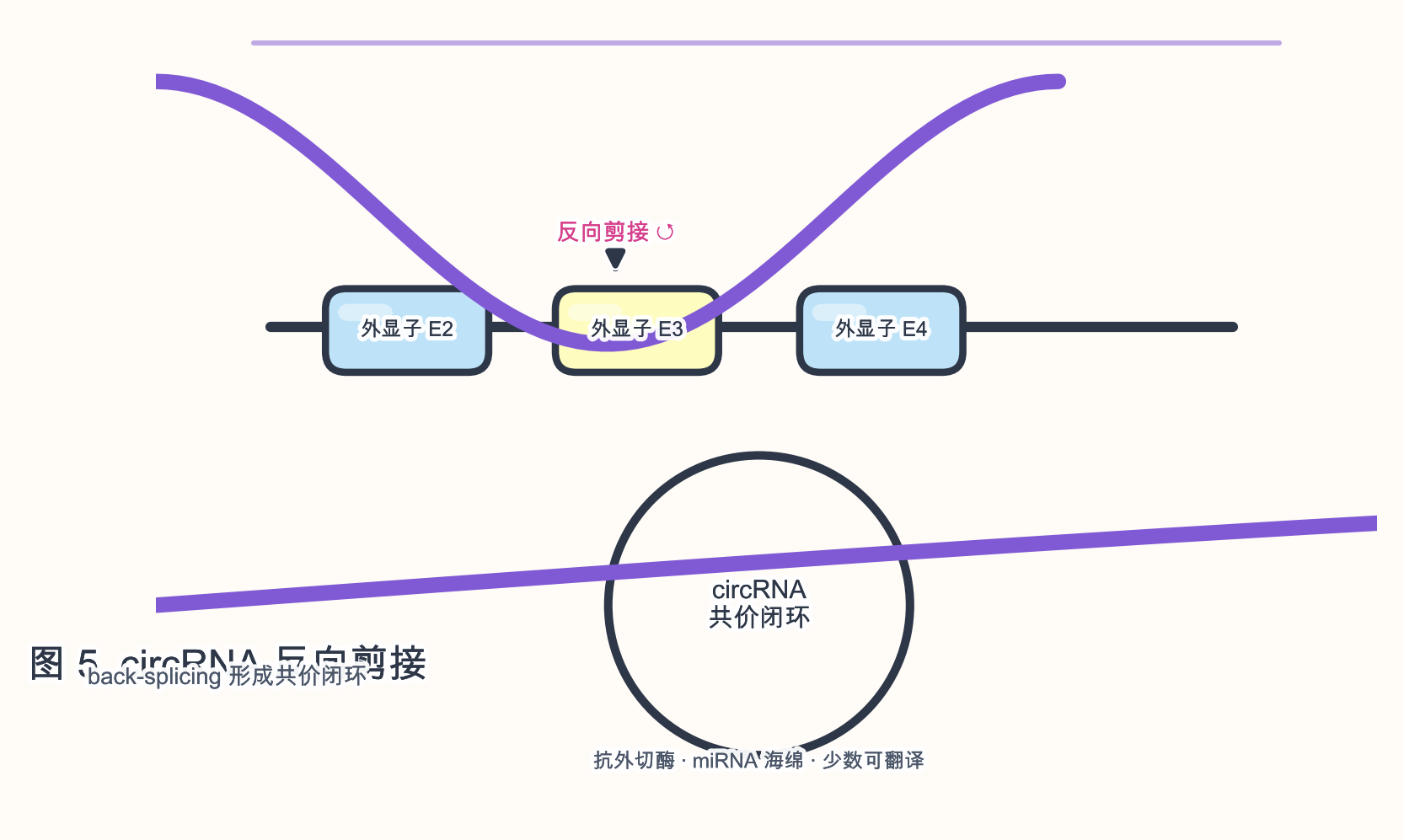
4.2 circRNA(环状 RNA)
生物学角色:circRNA 由 前 mRNA 经 反向剪接(back-splicing)形成 共价闭环,无 5’ 帽 与 3’ 尾,抗 外切酶 降解。
参与的生物学过程:miRNA 海绵(如 CDR1as/miR-7);RBP 隔离;IRES 介导的 翻译(少数);母源 circRNA 在生殖中的讨论。
序列与结构特征:
- 常由 外显子 环化,亦可含 内含子 序列;Alu 反向重复促进环化。
- RNase R 消化线性 RNA 后富集,是实验鉴定手段之一。
研究与应用:稳定 生物标志物;circRNA 疫苗 与 蛋白翻译 载体探索;与 lncRNA 共享 RNA-seq 分析流程但需专用工具(CIRI、find_circ 等)。
4.3 转录相关 ncRNA:eRNA、asRNA、PROMPT 等
| 类型 | 特征 | 功能概要 |
|---|---|---|
| eRNA(enhancer RNA) | 增强子 位点 Pol II 转录,常 非 polyA、短 | 增强子 染色质开放、三维互作 |
| asRNA(antisense RNA) | 与正义链 互补 | 转录干扰、染色质修饰、剪接 调控 |
| PROMPT / uaRNA | 启动子上游 | 启动子 近端调控、与 暂停 相关 |
| telomerase RNA(TERC) | H/ACA 样 | 端粒 延伸模板 |
段末注释:增强子(enhancer)为远端调控 启动子 活性的 DNA 元件;eRNA 是其活跃转录的 RNA 读出。
5. 特殊与情境型 ncRNA
5.1 tmRNA(转移–信使 RNA)
生物学角色:原核 tmRNA(SsrA)在 翻译停滞(如缺少终止密码子)时进入核糖体,恢复 翻译并标记 多肽 降解(反式翻译 rescue + 蛋白酶 标签)。
特征:~300 nt 量级,兼具 tRNA 样与 mRNA 样结构域;真核是否有完全等价物存在争议。
应用:细菌 应激 与 抗生素 耐受研究。
5.2 CRISPR RNA(crRNA)与 gRNA
生物学角色:原核 CRISPR–Cas 免疫系统中,crRNA 引导 Cas 核酸酶切割 外源 DNA/RNA。
特征:与 tracrRNA 或合成 单 guide RNA(sgRNA)配对;在基因编辑中 gRNA 为工程化 ncRNA。
应用:基因编辑、诊断(SHERLOCK、DETECTR)、基因敲低(Cas13)。
段末注释:CRISPR(clustered regularly interspaced short palindromic repeats)为成簇规律间隔短回文重复序列座。
5.3 病毒与人工 ncRNA
- 病毒 miRNA、RNA 基因组 非编码区(IRES、包装信号)。
- 适体(aptamer)、核酶、合成 shRNA(发夹经 Dicer 变为 siRNA 样)——在分类上常归入「工具 RNA」而非内源 ncRNA。
6. 衍生小 RNA 与「碎片化」读谱
高通量测序常检测到 tRNA/rRNA/snoRNA 的 片段,需与 miRNA 区分:
- tiRNA:应激下 血管紧张素 切割 tRNA 产生 5’/3’ tiRNA。
- tRF:多种 tRNA 来源小 RNA,参与 RISC 或非 RISC 通路。
- rsRNA:rRNA 片段,在 胞外囊泡 与免疫中有报道。
分析注意:文库构建 偏好会导致 rRNA 残留 dominate;生物信息 流程需 去 rRNA、miRBase/GtRNAdb 多重比对。
7. 横向对照:机制、长度与实验入口
表 1 主要 ncRNA 一览
| 类型 | 典型长度 | 主要定位 | 核心效应器/机器 | 典型功能 | 常用研究手段 |
|---|---|---|---|---|---|
| rRNA | kb 级 | 核糖体 | r-protein | 翻译催化 | 核糖体图谱、电镜 |
| tRNA | ~76–90 nt | 胞质/线粒体 | aaRS | 氨基酸转运 | tRNA-seq、氨酰化 assay |
| snRNA | ~100–300 nt | 核 | 剪接体 | pre-mRNA 剪接 | RNA-seq 剪接、CLIP |
| snoRNA | ~60–300 nt | 核仁 | fibrillarin/dyskerin | rRNA 修饰 | RiboMeth-seq、Northern |
| miRNA | ~22 nt | 胞质 | AGO/RISC | mRNA 抑制/切割 | qPCR、CLIP-seq、antimiR |
| siRNA | ~21 nt 双链 | 胞质 | AGO2 | 靶 mRNA 切割 | 转染、RNAi 药物 |
| piRNA | ~24–32 nt | 生殖细胞 | PIWI | TE 沉默 | piRNA-seq、ChIP |
| lncRNA | >200 nt–Mb | 核/质 | RBP、染色质复合体 | 多维调控 | RNA FISH、CRISPRi |
| circRNA | 变化 | 胞质为主 | RBP/miRNA | 海绵/翻译 | RNase R、circ 专用 caller |
| eRNA | 变化,常较短 | 核 | 转录机器 | 增强子活性 | GRO-seq、Hi-C |
表 2 siRNA、miRNA、shRNA 与内源小 RNA 边界
| 特征 | miRNA | siRNA(外源/设计) | shRNA(载体) |
|---|---|---|---|
| 来源 | 内源基因发夹 | 化学合成/体外 | Pol III/Pol II 表达发夹 |
| 配对 | 常部分,多靶 | 设计为高度特异 | 可设计为 siRNA 样 |
| 典型结果 | 微调表达 | 强 敲低 | 稳定 敲低 |
| 与 piRNA | 不同 AGO | 不同 AGO | 同左 |
8. 研究范式、数据库与 RNA 基础模型
8.1 实验与组学
- 表达:RNA-seq、small RNA-seq、单细胞 RNA-seq。
- 相互作用:CLIP 系列(HITS-CLIP、eCLIP)、RNA pull-down、SHAPE-MaP 结构 probing。
- 功能:CRISPR 敲除/激活、反义寡核苷酸、锁核酸 修饰寡核苷酸。
8.2 数据库与命名
| 资源 | 用途 |
|---|---|
| RNAcentral | 整合 ncRNA 序列 |
| Rfam | RNA 家族与协方差模型 |
| miRBase | miRNA 成熟体与发夹 |
| GENCODE | 人类 lncRNA 注释 |
| NONCODE | lncRNA 补充 |
| circBase | circRNA |
8.3 计算与 AI
RNA 基础模型(如 RNA-FM、RiNALMo)多在 混合 ncRNA 语料上预训练,嵌入 反映结构与功能而非单一 miRNA 类型;mRNA 专用模型(如 mRNA-FM)与 ncRNA 任务不可混用权重。详见 5003.大模型-架构-DNABERT-0.DNA-RNA预训练模型选型指南 中 ncRNA 语料说明。
9. 治疗与转化应用(按 RNA 类型)
| 领域 | 代表方向 |
|---|---|
| siRNA / ASO | 肝脏代谢病、神经系统(递送仍是瓶颈) |
| miRNA 模拟/抑制 | 纤维化、肿瘤(临床案例相对少) |
| antimiR | 病毒、心血管(早期临床探索) |
| lncRNA 靶向 ASO | 肿瘤 lncRNA(如 MALAT1 在研) |
| circRNA | 疫苗、蛋白 表达载体 |
| tRNA 工程 | 无义突变 抑制(tRNA 通读)、合成生物学 |
| CRISPR RNA | 基因编辑与 碱基编辑 标配组分 |
共同挑战:递送(LNP、GalNAc、胞内逃逸)、脱靶(种子区、免疫刺激 TLR)、化学修饰(2’-O-Me、PS 骨架)、耐久性与成本。
段末注释:ASO(antisense oligonucleotide)为反义寡核苷酸;LNP 为脂质纳米颗粒。
10. 与 siRNA 系列文档的关系
阅读本综述后,建议按主题进入:
| 文档 | 内容 |
|---|---|
siRNA-01-生物学机制与功能原理.md |
RISC、AGO2、链选择 |
siRNA-02-分子形式化学修饰与递送概览.md |
修饰与 LNP/GalNAc |
siRNA-05-设计指标脱靶与验证.md |
种子区、off-target |
ncRNA 全景帮助定位 siRNA 在「小 RNA 调控宇宙」中的位置:siRNA 是 RNAi 机器可装载的一种 双链底物形态,与内源 miRNA、endo-siRNA 共享 AGO 效应通路,但设计目标与配对规则更贴近「人造特异性 mRNA 切割器」。
小结
- ncRNA 覆盖从 催化型 rRNA 到 调控型 miRNA/lncRNA 的广泛谱系;单一长度阈值 不能代替功能分类。
- 管家 ncRNA(rRNA、tRNA、snRNA、snoRNA)支撑翻译与 RNA 成熟;小调控 RNA 多走 AGO/PIWI 通路;长 ncRNA 与 circRNA 更常重塑 染色质 与 网络。
- 序列(长度、修饰、配对)与 结构(发夹、环、RNP)共同决定功能;读谱时必须警惕 衍生片段 与 注释 混淆。
- 研究前沿 包括 单细胞 多维组学、RNA 结构组学、靶向 RNA 药物与 ncRNA 语言模型;应用上 siRNA/ASO 已商业化,lncRNA/circRNA 仍在快速演进。
- 配图 图 1–9 提供分类地图与代表性通路心智模型;细节实验设计请结合专题文档与原始文献。
延伸阅读(类型)
- ENCODE、FANTOM 非编码转录计划
- Rfam 14 / RNAcentral 发布说明
- 综述:Nature Reviews Molecular Cell Biology 系列 non-coding RNA 专题(按年份检索)